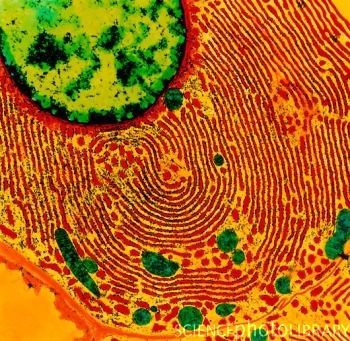
El retículo endoplasmatico es un organuelo que se encuentra alrededor de la célula y forma parte del sistema endomembranoso. Particular por que tiene dos tipos con funciones diferentes, el liso, el rugoso. Es un orgánulo formado por una serie de túbulos, sacos y vesículas rodeados de membrana e interconectados entre sí.
Retículo Endoplasmático Rugoso
El dominio rugoso del retículo endoplasmático se caracteriza por organizarse en una trama de túbulos alargados o sacos aplanados y apilados, más o menos regulares en su forma, con numerosos ribosomas asociados a sus membranas. La cantidad de ribosomas asociados a sus membranas condiciona la forma de este orgánulo, de tal manera que cuando el número de ribosomas asociados aumenta los túbulos se expanden adoptando la forma de cisternas aplanadas. Esta morfología es claramente visible en imágenes tomadas con el microscopio electrónico de las células secretoras de proteínas, las cuales tienen el retículo endoplasmático muy desarrollado.
La principal misión del retículo endoplasmático rugoso es la síntesis de proteínas que irán destinadas a diferentes lugares: el exterior celular, el interior de otros orgánulos que participan en la ruta vesicular, como los lisosomas, o que formarán parte integral de las membranas, tanto plasmática como de otros orgánulos de la ruta vesicular. Además, el retículo endoplasmático rugoso tiene que sintetizar proteínas para sí mismo, denominadas proteínas residentes. Las proteínas integrales de la membrana plasmática se sintetizan en el retículo endoplasmático. Hay que tener en cuenta que las proteínas sintetizadas en el retículo que van destinadas a orgánulos concretos de la ruta vesicular deben tener unas secuencias de aminoácidos o modificaciones específicas (actuarán como señales) para que cuando lleguen a la zona de reparto del aparato de Golgi sean reconocidas y dirigidas a sus compartimentos diana correspondientes.
Cualquier proteína que se secrete o que forme parte de los orgánulos o compartimentos de la ruta vesicular empieza su proceso de síntesis en el citosol pero terminará en el interior de una cisterna del retículo o formando parte de sus membranas. El proceso comienza con la unión de los ARNm, localizados en el citosol, uniéndose en primer lugar a una subunidad pequeña ribosomal y posteriormente a una subunidad grande ribosomal para comenzar la traducción. Lo primero que se traduce de estos ARNm es una secuencia inicial de nucleótidos a partir de la cual se sintetiza una cadena de unos 70 aminoácidos denominada péptido señal.
Una molécula conocida como SRP (sequence recognizing particule), que es una mezcla de 7 ARNs y 6 polipéptidos, reconoce al péptido señal y enlentece el proceso de traducción. El complejo formado por ribosoma, ARNm, péptido señal más SRP difunde por el citosol hasta chocar con las membranas del retículo endoplasmático, a las cuales se une gracias a la existencia de unreceptor de membrana que reconoce al SRP. Todo el complejo anterior interacciona con un translocador, que es una proteína integral que forma un canal por el cual penetra la cadena polipeptídica naciente hacia el interior de la cisterna del retículo endoplasmático.
El péptido señal queda unido al translocador mientras que el resto de la cadena que se va traduciendo y liberando hacia el interior. Una peptidasa presente en el retículo escinde el péptido señal del resto de la cadena de aminoácidos, quedando ésta libre en el interior. Una vez completada la síntesis, la cadena de aminoácidos adopta su conformación tridimensional y el ribosoma se libera de la membrana.
Si la proteína que se está sintetizando es una proteína integral de membrana no será liberada al interior del retículo. En estos casos las proteínas nacientes tienen cadenas de aminoácidos hidrófobos que cuando se traducen facilitan su inserción directamente entre los ácidos grasos de la membrana gracias a la acción del translocador. El proceso es muy complejo y diverso para los diferentes tipos de proteínas integrales puesto que en un receptor con siete cruces de membrana tienen que alternarse secuencias que han de insertarse en la membrana con otras que quedan bien en el lado citosólico o bien en el interior de la cisterna del retículo endoplasmático. Sólo en raras ocasiones el retículo importa proteínas que se sintetizan completamente en el citosol gracias a ciertos transportadores presentes en su membrana.
Las proteínas que se sintetizan en los ribosomas adosados a la membrana del retículo endoplasmático son modificadas conforme van siendo sintetizadas. a) Hay una glucosilación (N-glucosilación) de los aminoácidos asparragina. Éstos recibirán un complejo de 14 azúcares en su radical, que son transferidos desde un lípido embebido en la membrana denominado dolicol fosfato, perdiéndose algunos de estos azúcares en procesos posteriores. b) Se da hidroxilaciónsólo en algunas proteínas, sobre todo en aquellas que van a formar parte de la matriz extracelular. Aquí se hidroxilan los aminoácidos prolina y lisina, dando hidroxiprolina e hidroxilisina, que formarán parte del colágeno. c) Algunas proteínas asociadas a la membrana plasmática están unidas covalentemente a lípidos de la membrana, esta unión también se produce en este compartimento.
En el retículo endoplasmático se produce un control de la calidad de las proteínas sintetizadas, de modo que aquellas que tienen defectos son sacadas al citosol y eliminadas. Existen unas proteínas denominadas chaperonas que juegan un papel esencial en el plegamiento y maduración de las proteínas recién sintetizadas. Son también ellas las encargadas de detectar errores y marcar las proteínas defectuosas para su degradación. Otras proteínas con dominios tipo lectina, reconocen determinados azúcares y comprueban la adición correcta de glúcidos.
Formación y fusión de vesículas.
Como dijimos, las proteínas que se sintetizan en el retículo endoplasmático terminan en varios posibles destinos: en el exterior celular mediante un proceso de secreción, el interior o en la membrana de alguno de los compartimentos de la ruta vesicular como el aparato de Golgi, los endosomas o los lisosomas. Sin embargo, algunas tienen su función en el propio retículo endoplasmático, son las denominadasproteínas residentes. Hemos nombrado algunas como las chaperonas, ciertas glusosidasas, el receptor para el SRP, el propio translocados, etcétera. Para ser retenidas en el retículo deben poseer una secuencia de cuatro aminoácidos concrfetos localizados en el extremo carboxilo (-COOH).
Reticulo endoplasmatico liso
El retículo endoplásmico liso no presenta ribosomas. Sus funciones principales son la síntesis de lípidos de membrana, el almacenamiento de calcio y la detoxificación de drogas. Debido a esta última función, el retículo endoplásmico liso es muy abundante en hepatocitos y aumenta con la ingesta de sustancias tóxicas como el alcohol.
En células musculares lisas y estriadas encontramos una forma especializada de retículo endoplásmico liso conocida como retículo sarcoplásmico. El retículo sarcoplásmico es un importante almacén del calcio que se utiliza en el proceso de contracción muscular.
Síntesis lipídica
Las membranas del retículo endoplasmático liso producen la mayoría de los lípidos requeridos para la elaboración de las nuevas membranas de la célula, incluyendo glicerofosfolípidos y colesterol. Gran parte de la síntesis de los esfingolípidos se lleva a cabo en el aparato de Golgi, pero su estructura básica, la ceramida, se sintetiza también en el retículo. En realidad, en las membranas del retículo no se realizan todos los pasos
Detoxificación
Los hepatocitos, las células típicas del hígado, tienen un retículo endoplasmático liso muy desarrollado. En él se sintetizan las lipoproteínas que transportarán al colesterol y a otros lípidos al resto del organismo. En sus membranas se encuentran también enzimas, como la familia de proteínas P450, responsables de la eliminación de productos del metabolito potencialmente tóxicos, así como algunas toxinas liposolubles incorporadas durante la ingesta. La superficie de membrana del retículo se adapta a la cantidad de enzimas detoxificadoras sintetizadas, la cual depende a su vez de la cantidad de tóxicos presentes en el organismo.
Defosforilación de la glucosa-6 fosfato.
La glucosa se suele almacenar en forma de glucógeno, fundamentalmente en el hígado. Este órgano es el principal encargado de aportar glucosa a la sangre, gracias a la regulación llevada a cabo por las hormonas glucagón e insulina. La degradación del glucógeno produce glucosa-6-fosfato que no puede atravesar las membranas y por tanto no puede abandonar las células. La glucosa 6-fosfatasa se encarga de eliminar ese residuo fosfato, permitiendo que la glucosa sea transportada al exterior celular.
Reservorio intracelular de calcio
Las cisternas del retículo endoplasmático liso están también especializadas en el secuestro y almacenaje de calcio procedente del citosol. Este calcio puede salir de forma masiva en respuesta a señales extra o intracelulares gracias a cascadas de segundos mensajeros. Un ejemplo destacable es el retículo sarcoplásmico (nombre que recibe el retículo endoplasmático liso en las células musculares) que secuestra calcio gracias a una bomba de calcio presente en sus membranas. El secuestro y la salida de calcio desde el retículo sarcoplásmico se produce en cada ciclo de contracción de la célula muscular.
Algunas enfermedades estan asociadas con el reticulo endoplasmatico por ejemplo la diabetes y el mal de parkinson asociado indirectamente a este por lo que es importante tener en cuenta algunas caracteristicas y funcionamiento de este. De igual forma es difícil predecir a simple vista que tenemos algun mal asociado con esta organela aun asi es importante tener en cuenta algunos aspectos referentes a esta.
En conclusión el reticulo endoplasmatico asi como todas las organelas asociadas a la celula tienen gran importancia dentro de esta por que es un conjunto y como tal todas necesitan de cada una.
Referencias:
Referencias:
Anonimo. Sin año. Medicina Molecular. Reticulo Endoplasmatico. Documento PDF. Disponible en: http://www.medmol.es/glosario/102/
Bachmann,K.(1978) Biología para médicos:conceptos básicos para las facultades de medicina, farmacia y biología.Ed Reverte. 374 pags.
Ulrich Welsch, Johannes Sobotta(2009) Histología.España. Ed. Médica Panamericana.688 páginas
Marc Maillet.(2002). Biología celular. España. Ed. Elsevier. 537 páginas
Fleur . L. Strand. (1982). Fisiología humana: un enfoque hacia los mecanismos reguladores. España. Ed. Médica Panamericana. 694 páginas
Anonimo. Sin año. Atlas de Histología Vegetal y Animal. La célula. Reticulo Endoplasmatico. En linea. Disponible en: http://webs.uvigo.es/mmegias/5-celulas/5-reticulo.php
Universidad de Granma. Universidad Central de Las Villas.. Sin año. La web de la botanica. Reticulo Endoplasmatico. En linea. Disponible en:http://www.udg.co.cu/cmap/botanica/Reticulo_endoplasmatico.htm